 FEROMON PARFÜM
FEROMON PARFÜM FÉRFIAKNAK
FÉRFIAKNAK NŐKNEK
NŐKNEK ILLATMINTA
ILLATMINTA

Mi a feromon? Témakör: Érdekességek
Tartalomjegyzék:
Feromon
A feromon görög eredetű szó, a phero (φέρω = hord, visz) és hormon szavak egyesülésével keletkezett. Az élővilágban megtalálható olyan kémiai vegyületeket jelöl, melyek jelen vannak az állatvilágban és az emberekben is. A szerepük nagyon fontos, hiszen a feromonok hatására jön létre a kommunikáció legelső fajtái.
Befolyásolja, ezáltal irányítja a párkeresést és a szexuális érdeklődésre is hatással van. A feromon miatt válunk vonzóvá a másik fél szemében. Amikor nem tudod miért, de valakit ránézésre szimpatikusnak találsz, annak a feromon lehet az oka. Ezt használja ki a feromon parfüm is.
Így működik a feromon
A feromonok az apokrin mirigyekben fejlődnek és termelődnek. Levegővételkor a molekulák áthaladnak az orron, így aktivizálva az agyalapi mirigyünket. Az emberek orra körülbelül 10.000 szagot tud megkülönböztetni. Egy részük a szaglósejtekből álló szaglóhám előtt hömpölyög.
Kutatási eredmény – Feromonokat találtak az emberi apokrin mirigyekben
Az apokrin mirigyek módosult verejtékmirigyek, amelyek a pubertás utáni szőrt kifejlő területeken – a hónaljban és a szeméremkörben – találhatók és funkciójuk túlnyomórészt a szagkiválasztás. Ezeken a területeken (az egyébként nagyrészt meztelen testünkön) a szőrzet speciális fejlődése elősegíti a szaganyagok szétterjedését ivarérett emberekben.
A szőrzet meleg környezetet biztosít, ahol a kommenzális baktériumok hatása elpárologtatja az apokrin mirigyekből felszabaduló prekurzor molekulákat. [12]
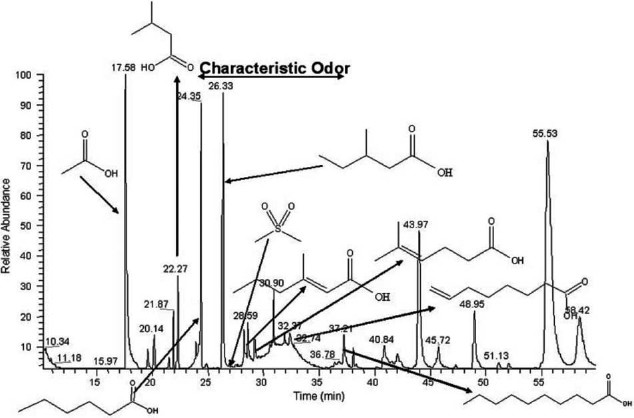
Emberi feromon izzadság gázkromatogramja
Feromon fajták
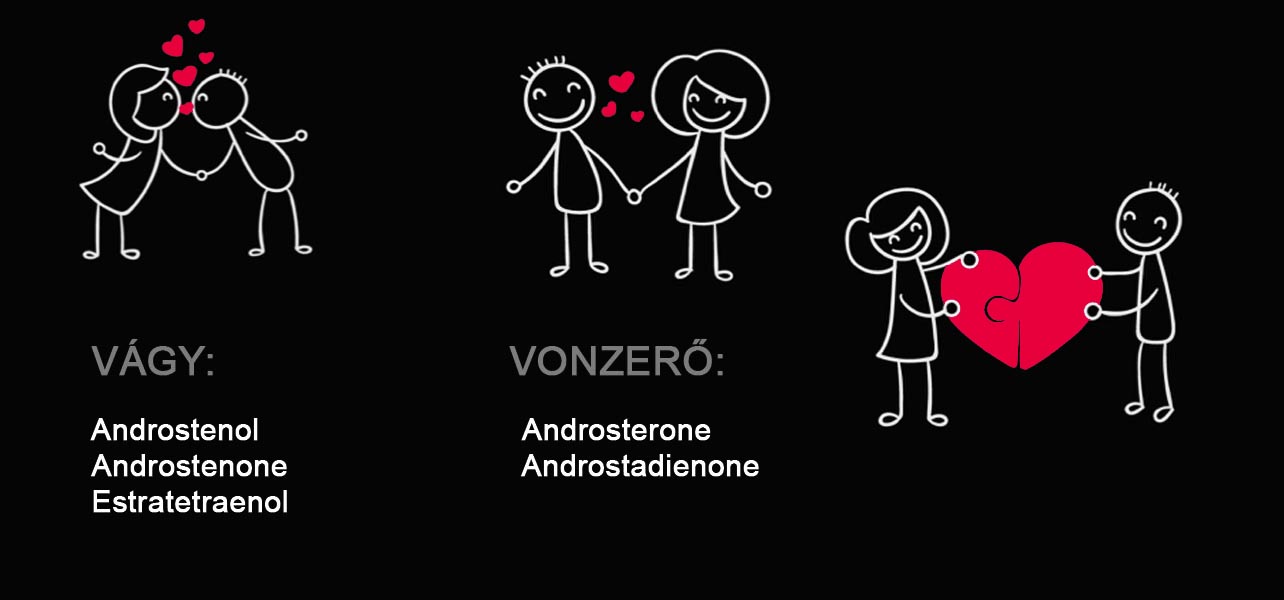
Androstenol:
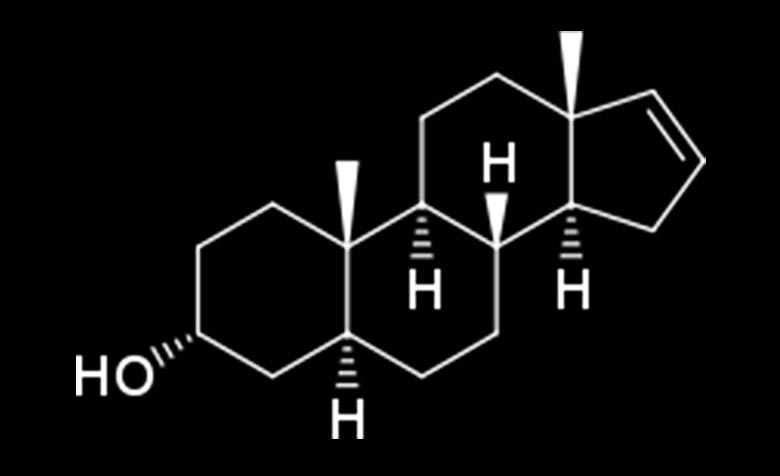
Stresszoldó hatást vált ki, kutatások bizonyítják, hogy hatással van a viselkedésre, ami azt jelenti, hogy idegen emberekkel is könnyebben beszélgetést kezdeményezünk, megkönnyíti a kommunikáció elsődleges formáját.
Androstenone:
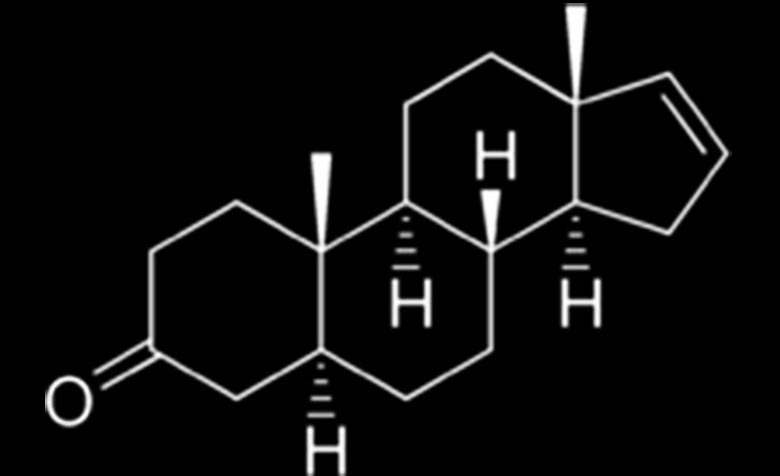
Tanulmányok során a legelső felismert feromon az androstenon. Először férfiak nyálában ismerték fel, a szervezetük termeli és ezáltal a női nemet vonzza. A szervezet által termelt androstenont a nők csupán 0,2 százaléka érzékeli. Az androstenon feromont viselő férfi domináns, meghatározó benyomást kelt a nőben.
Androsterone:
Az emberi hónaljban és a bőrben mutatták ki ezt a fajta feromont. A biztonságérzetet növeli a női nemben, határozott, irányító férfi típus érzését közvetíti. A nőkre nagy hatással van ez a feromon, hiszen ezáltal kap megerősítést, hogy megbízható férfi társaságában van.
Androstadienone:
Az elengedhetetlen „szerelmes feromon”. Kellemes, megnyugtató hatást fejt ki mely jótékony hatással van a nőkre. Cselekvésre ösztönzi őket és meghitt boldog pillanatokat okozhat ez a feromon.
Estratetraenol:
Erre a feromonra reagálnak a nők a legerősebben. Nagyobb társaságban is képes kifejteni a hatását. Keresni fogják a társaságod, keresik a szemkontaktust, a beszélgetés során élvezni fogják a társaságod, kellemes partnert találnak a személyedbe. AndrostaDIENOL :
Tudtad? A Feromonok az állatvilágban is megtalálhatóak, egyes rovarok olyan anyagot képesek kibocsátani, amellyel magukhoz vonzzák hasonló fajú társaikat.
Így hat a feromon a szerelemben
A párkeresés során a vonzalom kialakulásáért a feromon a felelős. Nézzük meg ezt részletesebben, tudományos módon.
A szerelem neurobiológiai folyamatai mélyen beágyazódnak az emberi agy szocioérzelmi rendszerébe, ahol kulcsszerepet játszanak a kémiai jelzőmolekulák, köztük a feromonok. Ezek az illékony vegyületek képesek észrevétlenül befolyásolni a vonzalmat, a szexuális vonzódást és a kötődési folyamatokat. A feromonok hatásmechanizmusa a szaglóhámon (MOB) keresztül indul, és specifikus agyi területeket aktivál, amelyek közvetlenül kapcsolódnak a szerelem élményéhez. [21]
A feromonok érzékelése és feldolgozása
Röviden, mert erre később részletesen kitérünk; az egyes emberi feromonok receptorai az orrüreg hátsó részén található VNO-ban helyezkednek el. Mások pedig a szaglóhagymában (MOB) [0] Ezek a receptorok a G-fehérje kapcsolt jelátviteli rendszeren keresztül közvetítik a jelet a limbikus rendszer irányába. A szaglási ingerület először az accessory olfactory bulb (AOB) régióban kerül feldolgozásra, majd továbbhalad az amygdala, a hypothalamus és a prefrontális kéreg irányába. [21] [13]
Fontos idegi célterületek:
- Amygdala: érzelmi válaszok feldolgozása
- Hypothalamus: hormonális reakciók és szexuális motiváció
- Orbitofrontális kéreg: tudatos érzelmi élmény, vonzalom
A szerelem neurokémiai komponensei
A feromonok szerepe nem elszigetelt, hanem szinergikus más neurokémiai anyagokkal, amelyek a romantikus kötődés élményét közvetítik. [14] A feromonok által kiváltott ingerek hatására fokozódik bizonyos neurotranszmitterek és hormonok szintje, mint például [15] :
- Dopamin: a jutalmazási és motivációs rendszer központi vegyülete
- Oxitocin: kötődés, bizalom és intimitás elősegítő hormon
- Vazopresszin: hosszú távú partnerkötődést támogató hormon
- Noradrenalin: izgalmi állapot, eufória, figyelemfókuszálás
Ezek a molekulák párhuzamosan aktiválódnak a VNO és a MOB által közvetített feromonjelek hatására, erősítve a romantikus kötődést és szexuális vonzalmat.
A tudattalan vonzalom
Az emberi test által kibocsátott feromon mintázatok a genetikai különbségeken – főként a fő hisztokompatibilitási komplex (MHC) variációin – alapulnak. Az eltérő MHC-profilokat mutató személyek között erősebb lehet a feromonok által közvetített vonzalom. Ez biológiailag előnyös, mivel az utódok immunrendszerének sokszínűségét növeli.
Az emberi szerelem mögött komplex neurobiológiai és neurokémiai folyamatok húzódnak, amelyekben a feromonok kiemelt jelentőséggel bírnak. Ezek a szagtalan kémiai jelek nemcsak a vonzalmat és szexuális érdeklődést befolyásolják, hanem a hosszú távú kötődés kialakulását is támogatják. A feromonok, a VNO és a MOB által közvetített jelek és az agyi válaszreakciók együttesen alkotják azt a biológiai hátteret, amely lehetővé teszi a szerelem tudattalan, de mélyen emberi élményét. [16]
Feromon érzékelése
Minden egyes sejtnek 10-20 csillója van, amik egyesülnek az orrüreg hámbélését borító nyálkahártyával. Azok a molekulák, amelyek oldódnak a nyálkában, és a csillókat ingerelve idegingerületet alkotnak. Az axonok összeolvadva a szaglógumóba továbbítják az adatot, ahol a feldolgozás elkezdődik. Ezután a szagjelek a középső szaglóterületbe és az amigdalában levő oldalsó szaglóterületbe érnek. Ezeken a részeken folyik a szaglással kapcsolatos agytevékenység legnagyobb része.
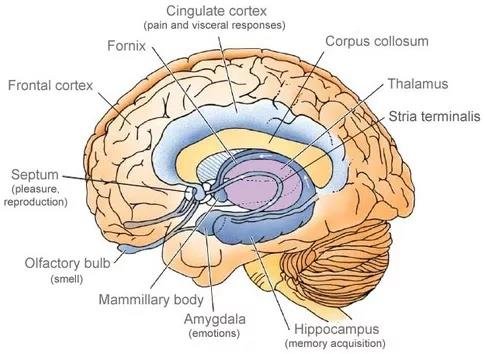
A feromonokat nem tudatosan érzékeljük, mégis befolyásolják az utódnemzéshez kapcsolódó folyamatokat, hatással van viselkedésünkre. Például, amikor nem tudod miért, de vonzónak találsz valakit, annak a feromon (is) az oka.
A feromonok jelenlétét érzékeli az orrban jelenlévő vomeronazális szerv (VNO) továbbá a (MOB) és információt közvetít az agyunknak. Az emberi vomeronazális szerv aktív szerepet játszik a feromonok érzékelésében, és közvetlen kapcsolatban áll az agy érzelmi és hormonális központjaival.
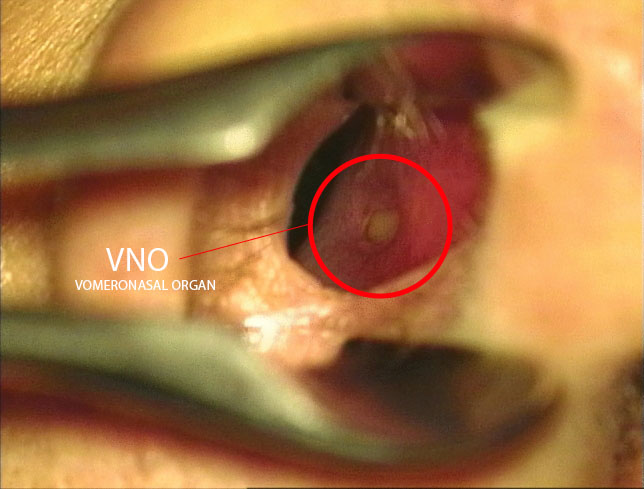
A vomeronazális szerv (VNO) szerepe az emberi feromon érzékelésben
A VNO működésének és a feromonok hatásainak kutatása mélyebb betekintést nyújt az emberi viselkedés és szociális interakciók neurobiológiai alapjaiba. Az emberi vomeronazális szerv egy specializált kémiai érzékelő rendszer, amely képes feromonok detektálására, és amely közvetlen kapcsolatban áll az agy szociális és hormonális szabályozásért felelős régióival.
Bár a VNO szerepe sokáig kérdéses volt és rengeteg weboldalon még a régi pontatlan kutatásokat lehet olvasni. Van olyan vita oldal, ahol 1931-es tanulmányra hivatkoznak, amikor még nem létezett olyan technológia, amivel a mai kutatásokat meg lehetett erősíteni. Szerencsére a növekvő számú neurobiológiai és molekuláris adat alátámasztja a VNO funkcionális jelenlétét és hatását az emberi viselkedésre, azonban az is kiderült, hogy csak bizonyos feromonokat érzékel és a továbbiakban a MOB (szaglóhagyma) veszi át a a VNO szerepét. Erről részletesebben lentebb beszélünk majd egy Magyar kutatók részvételével végzett kutatásban.
Neurobiológiai mechanizmusok és agyi folyamatok
A vomeronazális szerv (VNO), más néven Jacobson-szerv, egy kémiai érzékelő rendszer része, amely az emberi orrüreg alsó részében helyezkedik el. Bár a tudományos közösség egy része vitatja annak funkcionális szerepét az emberben, számos anatómiai, molekuláris és neurobiológiai bizonyíték támogatja azt az elképzelést, hogy a VNO aktív szerepet játszik a feromonok észlelésében, és képes befolyásolni viselkedést és endokrin válaszokat. Annyi mára bizonyos, hogy a VNO és a MOB együtt érzékeli a különböző feromonokat, hiszen rengeteg fajta ilyen illékony vegyület létezik.
Beszéljünk egy kicsit az emberi VNO szerkezetéről, feromon érzékelő képességeiről, valamint az érzékelési folyamat során bekövetkező idegrendszeri mechanizmusokról, különös tekintettel a központi idegrendszeri pályákra és az agyi területekre. [1] [6]
A VNO anatómiája és működése
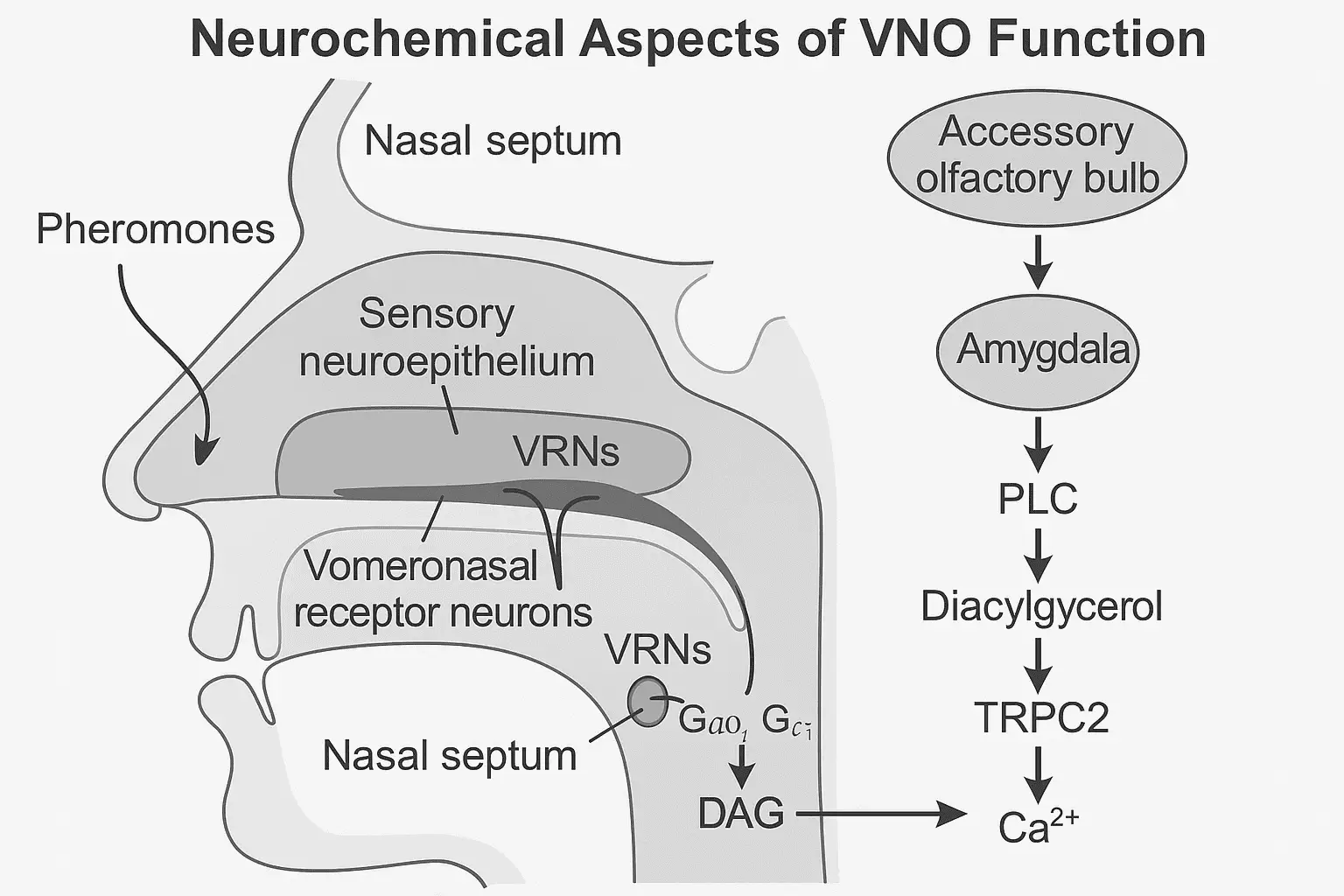
A VNO az emberi orrsövény ventromediális részében található, a nasopalatális csatorna környezetében. Mikroszkópos vizsgálatok igazolták, hogy ez a szerv tartalmaz szenzoros neuroepitéliumot, amely specializált receptorsejteket foglal magába. Ezek a sejtek képesek kémiai jelzések, különösen feromonok detektálására. A VNO-receptorok közé tartoznak a V1R és V2R típusú G-fehérjéhez kapcsolt receptorok, amelyek közvetlenül kapcsolódnak másodlagos hírvivő rendszerekhez, például a foszfolipáz C és a diacilglicerol útvonalhoz. [2] [7]

A vomeronasalis receptor géncsalád, valamint a V1R és V2R receptorok szerkezete.
Feromon érzékelés mechanizmusa
A VNO receptorai szelektíven reagálnak bizonyos szerves vegyületekre, például szteroid eredetű feromonokra, amelyek az emberi izzadságban, nyálban és egyéb testváladékokban találhatók. Ezek a molekulák kötődnek a receptorfehérjékhez, aktiválva intracelluláris jelátviteli utakat, amelyek elektromos impulzusokat generálnak.
A jelátvitel során keletkezett ingerület a vomeronazális idegen (n. vomeronasalis) keresztül a segítő olfaktorikus rész (accessory olfactory bulb, AOB) felé halad, majd innen az amygdala, a hypothalamus, és a limbikus rendszer egyéb régiói felé. Ezek az agyterületek kulcsszerepet játszanak az érzelem feldolgozásban, társas viselkedésben és hormonális szabályozásban. [3] [8]
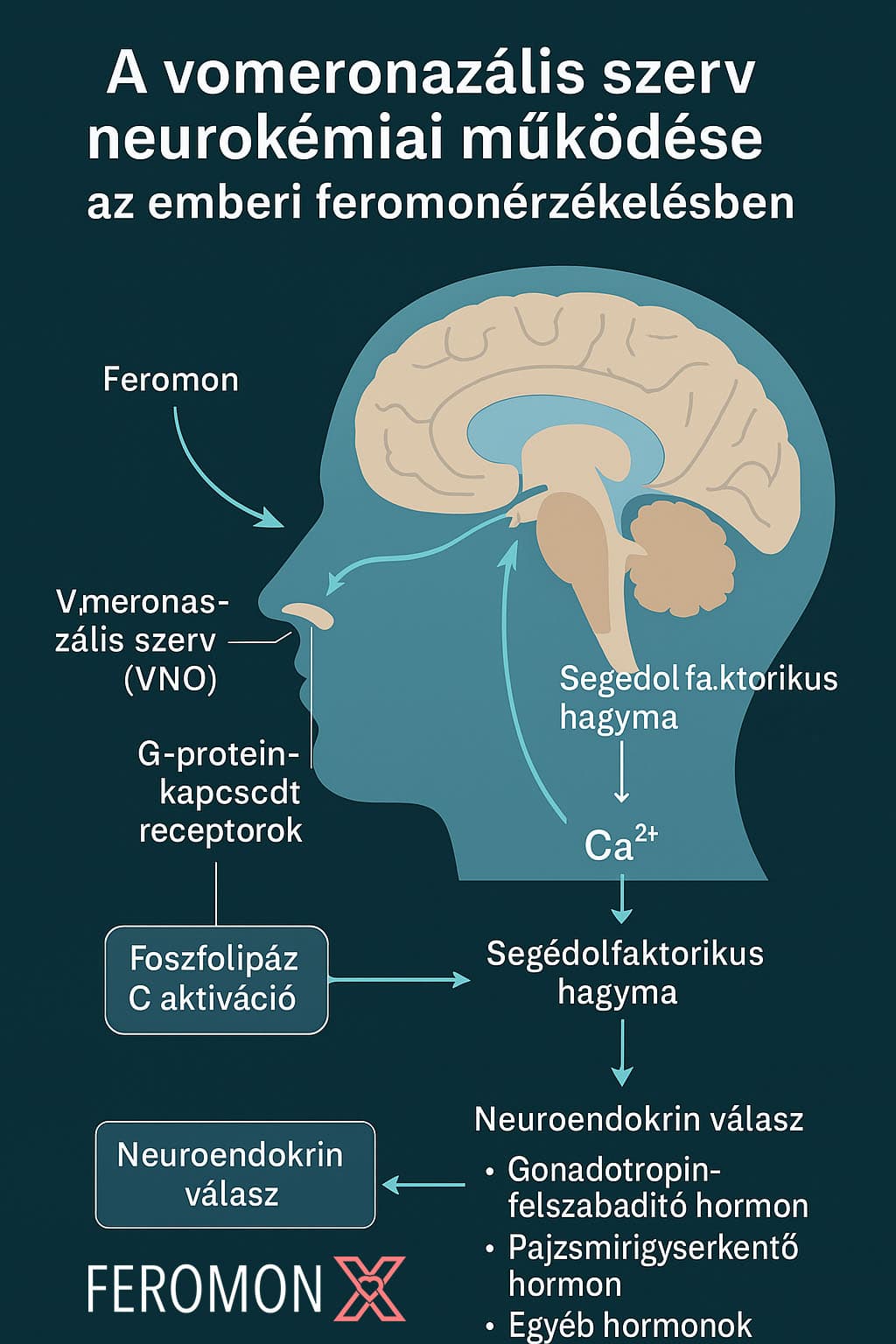
Agyi feldolgozás és viselkedési hatások
Az amygdala aktivációja révén a feromon jelek tudattalanul befolyásolhatják az érzelmi állapotot, szexuális vonzalmat és társas preferenciákat. A hypothalamus, mint a hormonális szabályozás központja, neuroendokrin válaszokat vált ki, például luteinizáló hormon (LH) vagy tesztoszteron szint változást. Ezek a válaszok nem tudatosulnak, mégis képesek módosítani az emberi viselkedést és fiziológiát, különösen a szociális kontextusban. [4] [9]
Klinikai és kísérletes megfigyelések
Funkcionális képalkotó eljárásokkal (pl. fMRI, PET) végzett vizsgálatok azt mutatták, hogy bizonyos illatanyagok – különösen azok, amelyeket feromon vegyületként jellemeznek – fokozott aktivitást váltanak ki az amygdalában és a hypothalamusban. Emellett egyes tanulmányok humán VNO régióban VNO-receptorokat kódoló gének expresszióját is igazolták, alátámasztva e szerv szenzoros potenciálját. [5]
A vomeronazális szerv neurokémiai működése az emberi feromon érzékelésben
Az emberi vomeronazális szerv (VNO) kémiai jelek, különösen feromonok érzékelésére specializálódott struktúra. Neurokémiai működése komplex receptor-közvetített jelátvitelen alapul, amelyben G-fehérje-kapcsolt receptorok, másodlagos hírvivők és központi idegrendszeri pályák vesznek részt.
Feromon receptorok és jeltovábbító mechanizmusok
A VNO receptorneuronjai G-protein kapcsolt receptorokat (GPCR) expresszálnak, főként a V1R és V2R típusokat. Emberben ezekhez társuló G-protein alegységek közül kiemelkedik a Gαi2 és Gαo, amelyek kulcsszerepet játszanak a jelátvitel inicializálásában. A feromon kötődése a receptorhoz aktiválja a G-fehérjét, ami elindítja a foszfolipáz C (PLC) enzim működését. [10]
A MOB további feromonokat érzékel, olyanokat, amelyeket a VNO esetleg nem képes. [21]
A PLC lebontja a foszfatidil-inozitol-4,5-biszfoszfátot (PIP2) inozitol-1,4,5-triszfoszfátra (IP3) és diacilglicerolra (DAG). Ez a másodlagos hírvivő rendszer intracelluláris kalcium felszabadítást okoz, ami depolarizációt és akciós potenciált generál. Az így kialakult ingerület továbbhalad a VNO és MOB idegen keresztül az agy felé. [11]
Érthetőbb, nem szakmai nyelven:
Ekkor történik meg az első benyomás
Ez alapján már tudat alatt eldöntöttük, hogy vonzódunk e ahhoz akire épp ránézünk vagy beszélünk.
Ha szerelmes vagy a párod testének/bőrének az illatát vonzónak találod. Az illatot nem tudod meghatározni, csak azt tudod, hogy jó és szívesen vagy a közelében. Több testi kontaktust igényelsz és kezdeményezel is. Ha napközben távol vagytok egymástól, az orrodban tovább örzöd az illatát amit bármikor feltudsz idézni. Jó érzés és boldogság jár át az illat hatására. Ha hosszútávú, stabil párkapcsolatban élsz, akkor segít fenntartani a vonzalmat és felpezsdíti a hétköznapokat is.
Számos kutatás és vakteszt eredménye azt mutatja, hogy 10 különböző ember illatából is képesek a szerelmesek felismerni a saját párjuk illatát.

A testszag olyan mint az ujjlenyomat, mindenkinek egyedi. Befolyásolja a mindennapi életvitel, az étkezési szokások sőt az is, hogy milyen típusú (száraz vagy zsíros) a bőröd.
Ha azt tapasztalod, hogy a kiszemelted, tökéletesen néz ki, kedves, jó vele beszélgetni, még sincs meg az a bizonyos szikra az a plusz, annak az az oka, hogy hiányzik a feromon. Ez miatt nem alakul ki szexuális vonzódás és legjobb esetben is csak jó barátok lesztek.
Bizonyíték a feromonra
Az emberi feromonok létezésének tudományos bizonyítékai
Áttekintés a humán kémiai kommunikáció kutatási eredményeiről. A feromonok olyan kémiai jelek, amelyeket egy egyed választ ki, és amelyek viselkedési vagy élettani válaszokat váltanak ki egy másik egyedben, ugyanazon fajon belül. Az állatvilágban jól dokumentált a feromonok szerepe, az emberi feromonok létezése is bizonyítást nyert különböző neurológiai, pszichológiai és biokémiai kutatások révén. [21] Mi most csak az emberi feromonokra fókuszálunk, azok hatásmechanizmusát bemutatva.
Molekuláris azonosítás
Kutatók több olyan vegyületet is azonosítottak, amelyeket az emberi test természetes módon választ ki, és amelyek kémiai jelként működnek. Különösen az apokrin mirigyekből származó szteroidok (pl. androstenone, androstenol) mutattak ki feromon hatásokat. [17]
Kulcsvegyületek:
- Androsztadienon (a férfi verejtékben található; nőkben hangulat- és hormonváltozásokat okoz)
- Estratetraenol (női eredetű illékony vegyület, férfiak viselkedését és észlelését befolyásolja)
Neurológiai válaszreakciók
A feromon vegyületek belélegzése közvetlenül aktiválja a hipotalamusz egyes területeit, ez az agy hormonális és viselkedési válaszaiért felelős központja. [17] Az androstenone például nőknél fokozott agyi aktivitást váltott ki a preoptikus és ventromediális hipotalamuszban, amelyek szerepet játszanak a szexuális válaszreakciókban. [18]
Viselkedési és szociálpszichológiai hatások
Az emberi feromonok képesek befolyásolni a társas viselkedést, hangulatot és párválasztást. Kísérletek során kimutatták, hogy az androstenone jelenléte nőknél pozitívabb hangulatot és megnövekedett figyelmet váltott ki a férfiak felé. Egy másik vizsgálat szerint a női szagnyomok (androstenol) hatására férfiak jobban ítélték meg a női arcokat. [19]
Az emberi feromonok nemcsak léteznek, hanem specifikus molekuláris, idegrendszeri és viselkedési hatásokat fejtenek ki. A különféle szteroid eredetű vegyületek az emberi szervezet által termelődnek, és bizonyíthatóan befolyásolják a társas és szexuális viselkedést, mindezt idegi aktiváció és hormonális szabályozás révén. A bizonyítékok sokrétűsége alátámasztja, hogy a feromonális kommunikáció az emberi viselkedés szerves része. [20]
A tudományos tanulmányok alapján biztosan állítható, hogy a férfiak és a nők is termelnek feromont, de a mai felgyorsult világ hatására ezek elkopnak. Őseinknek kevesebb lehetőségük volt a tisztálkodásra, ezáltal jobban megőrizték a bőrük természetes illatát.

Ma már ez nem lehetséges, hiszen mindennap fürdünk, a ruháinkat rendszeresen mossuk és különféle illatanyag tartalmú kozmetikumokat használunk a testünk különböző részeire mindennap. Ez vezet ahhoz, hogy a saját testünk által termelt feromon kevésnek bizonyul ahhoz, hogy a hatását kifejtse.
Először 1974-ben ismerték fel a feromont az emberi verejtékben az androsztenolt. A kutatást végző tudós kijelentette, hogy megtalálta az első olyan emberi feromont, ami a szexualitást és a vonzalmat befolyásolja.
A Madison egyetem kutatásai során a selyemmajmokon tanulmányozták a feromon működését és hatását. Ezáltal kiderült, hogy az állatok által kibocsátott feromonok hatással vannak a fajtársak agyműködésére. Viselkedésbeli változást idézett elő a hím és a nőstény majmoknál is. A zárkózott és visszahúzódó állatok viselkedése megváltozott, nyitottabbak lettek fajtársaikra, szaporodásra való hajlandóságuk megnőtt. A magányos egyedek pedig csoportosulni kezdtek.
David Berliner vizsgálatai alapján megtudhattuk, hogy az emberek felszabadultabbak és közvetlenebbek lesznek egymás társaságban ha a levegőben feromon van jelen. Az emberek a munkahelyükön jobban teljesítettek, oldottabbak voltak egymással, így a teljesítményük is növekedett.
Áttörés az emberi feromonok érzékelése kutatásában – magyar kutatók részvételével
A Nemzeti Biotechnológiai Információs Központ (NIH) által közölt kutatás szerint:
“Az eredmények együttesen alátámasztják azt a hipotézist, hogy az emberek, számos más emlőshöz hasonlóan, képesek a feromonjelek feldolgozására a szaglónyálkahártyán keresztül.”
forrás / kutatás: Feromon jelátvitel emberekben Kutatók: Dr. Gulyás Balázs, Ivanka Savic, Ebba Hedén‐Blomqvist, Hans Berglund – Stockholmi Agykutató Intézet, Klinikai Idegtudományi Osztály, Svédország PMCID: PMC6870699
A kutatás eredményének könnyebb megértéséhez szükség van néhány fogalom magyarázatára.
Mi a feromon?
Karlson és Luscher [ 1959 ] eredeti definíciója szerint a feromonok „levegőben terjedő kémiai jelek, amelyeket egy egyed bocsát ki a környezetbe, és amelyek befolyásolják ugyanazon faj többi egyedének fiziológiáját és viselkedését”. Mind az illékony, mind a nem illékony vegyületek feromonként működhetnek. Ezek a kémiai jelek információt nyújtanak a nemről és a reproduktív állapotról, és közvetítik a társadalmi és szexuális viselkedést, valamint a neuroendokrin változásokat. Az emlősökben a feromonok kimutatásának fő érzékszerve a vomeronazális szerv (VNO).
Mi a VNO?
A VNO egy vak végű tasakban található az orrsövényben, és egy porcos tok veszi körül, amely egy kivezetőcsövön keresztül nyílik az orrüreg aljába [Meredith, 1991 ; Monti‐Bloch et al., 1998 ].
A VNO egyértelműen eltávolodik a szaglónyálkahártyától, amely az orrüreg tetején található. Állatkísérletes adatok azt mutatják, hogy az orrüregben található fő szaglóhám szaglóérzékelő neuronjainak axonjai a fő szaglóhagymába (MOB) vetülnek ki, míg a VNO neuronjainak axonjai a járulékos szaglóhagymába (AOB) vetülnek ki. A jelenlegi nézet az, hogy az illékony szaganyagokat a szaglóhám érzékeli, míg a feromonokat a VNO-n keresztül [Keverne, 2007 ]. Az állatoknál a feromonjelzés által befolyásolt viselkedéseket, mint például a szexuális viselkedést, az agressziót és a terhességi blokkot, jellemzően a VNO által közvetített folyamatoknak tulajdonítják [Keverne, 2007 ].
Ha jobban megvizsgáljuk a kutatásokat, akkor a legtöbb olyan témájú kutatás, amely azt az eredményt közölte, hogy az ember nem képes a feromonokat érzékelni, egytől-egyig a VNO-n keresztüli vizsgálatot részesített előnyben.
Miért? Mert a kutatók (hivatalosan) állatokon tesztel(het)nek, és az állatokban (is) megtalálható VNO-t hasonlították össze az emberi orrban található VNO-val. Állatokon végzett tesztek alatt azt értjük, hogy például egér orrából kioperálják a VNO szervet és úgy vizsgálják utána, hogy az egyed érzékeli-e VNO nélkül a feromonokat (egyébként igen, VNO nélkül is érzékeli). Mivel embereken ilyen kisérleti operációkat nem alkalmaznak – hivatalosan legalábbis biztos nem.
A mostani kutatások azt bizonyítják, hogy ez a fajta megközelítés helytelen volt. Nem azt vizsgálták, hogy az ember hogyan érzékeli a feromonokat, hanem, hogy a VNO-n keresztül érzékeli-e a feromonokat és a kutatók arra jutottak, hogy nem.
De ettől még az ember érzékeli a feromonokat, csak kicsit másképp.
Korábbi kutatásokból idézetek, melyek a fél internetet és az AI ügynököket is félrevezették:
- Bár az embereknek is van VNO-juk, számos jel utal arra, hogy születés után nem funkcionális [Trotier et al., 2000 ]. Először is, az emberi VNO hámja jobban hasonlít a légzőhámra, mint a funkcionális VNO-val rendelkező fajoknál található VNO neuroepitheliumra [Witt et al., 2002 ].
- Másodszor, az olfaktorikus marker fehérje (OMP), amely az érett VNO neuronok megbízható markere, nem expresszálódik az emberi VNO-ban [Dennis et al., 2004 ].
- Harmadszor, a feromon jelátvitelhez szükséges TRCP2 ioncsatornákat kódoló gének pszeudogénként működnek az emberekben [Zufall et al., 2002 ].
- Továbbá az AOB részt vesz az emberekben, míg az összes többi funkcionális VNO-val rendelkező fajnál a feromonjelek feldolgozásáért felelős közvetítő struktúra [Dennis et al., 2004 ].
Ennek következtében a feromonjelátvitelt régóta megkérdőjelezik az emberekben. De végre tisztázódni látszik a helyzet egy másfajta kutatási logika alapján, mert ugye nem mindegy, hogy:
Az ember képes-e érzékelni a feromonokat a VNO-n keresztül – amire a válasz röviden egyesek szerint; nem vagy nem mindegyiket, vagy megkérdőjelezhető – ezeket a rövid válaszokat kiemelve a teljes kutatás kontextusából a mainstream média lehozza, majd erre mindenki azt gondolja, hogy az ember nem érzékeli a feromonokat.
vagy
Hogyan érzékeli az ember a feromonokat? – kérdést tesszük fel és kutatjuk.
Érzékelhető a két nézőpont közötti különbség. Miért érdekes ez? Mert nem az a kérdés, hogy az ember érzékeli-e feromonokat, mert az neurobiológiai tény évtizedek óta – , hanem pontosan hogyan érzékeli és milyen hatással van ez a viselkedésére.
Tehát, mit mondtak eddig a kutatók és mit közölt a kattintás hajhász média?
“Mivel a VNO szerv felnőtt emberekben nem működik, a feromonok viselkedésünkre gyakorolt hatása erősen megkérdőjelezhető.”
Ezt a nézetet szerencsére ellentmondják a feromonok emberi fiziológiára és viselkedésre gyakorolt hatásával kapcsolatos egyre növekvő kutatások, érvek és bizonyítékok.
- Az egyik ilyen érv az, hogy a női szobatársak menstruációs ciklusainak jól ismert szinkronizációját az izzadság közvetíti, amely feromonszerű vegyületeket tartalmaz [Stern és McClintock, 1998 ].
- Egy másik érv az, hogy a férfi izzadságot szagló nők a luteinhormonjuk pulzatilitását az ovuláció elősegítése érdekében megváltoztatják [Preti et al., 2003 ].
- Továbbá, a feromonszerű vegyületek, a 4,16-androsztadien-3-on (AND) ösztra-1,3,5(10),16-tetraén-3-ol (EST) több egymást követő kísérletben is kimutatták, hogy befolyásolják a hangulatot és az izgalmi szintet [Bensafi et al., 2004 ; Jacob et al., 2001 ; Jacob és McClintock, 2000 ; Lundstrom és Olsson, 2005 ].
- Végül a PET és az fMRI kimutatta, hogy ezen vegyületek szaglása aktiválja az emberi agy specifikus, reprodukcióval kapcsolatos neuronális áramköreit, nem és szexuális irányultság szerint differenciált módon [Berglund et al., 2006 ; Savic, 2002 ; Savic et al., 2001 , 2005 ; Sobel et al., 1999 ].
Tehát, ha az emberek nem rendelkeznek működőképes VNO-val, akkor az a kérdés, hogyan lehetséges, hogy képesek feromon jeleket detektálni?
Ahogy korábbi tanulmányokban is tárgyaltuk, a megfigyelt agyi aktivációk közvetítődhetnek 3 módon keresztül [Savic et al., 2001 ]:
- VNO
- szagló nyálkahártya
- vér
Ezen alternatívák szétválasztása és a szagló nyálkahártya hatásának vizsgálata érdekében az új kutatásban egy másik megközelítést használtak, PET aktivációs vizsgálatokat végeztek az EST és az AND szaglása során 12 krónikus anozmiában szenvedő heteroszexuális férfin.
Az anozmia az orrpolipokon alapult, amely akadályozza a jelátvitelt a szaglónyálkahártyán keresztül, és nem befolyásolhatja a VNO-t.
A kutatók abból indultak ki, hogy feltételezték, hogy ha egy VNO által közvetített útvonal működőképes, az EST szaglása hipotalamusz aktivációt eredményezne az anozmiás férfiaknál, ahogyan az egészséges heteroszexuális férfiaknál is történt [Berglund et al., 2006 ].
Az agyi aktiváció hiánya ezzel szemben a szaglónyálkahártyán keresztüli jelátvitelt részesítené előnyben.
Annak biztosítása érdekében, hogy a funkcionális károsodás a szagló nyálkahártyára korlátozódjon, az aktivációs kísérletek magukban foglalták a vanillin (VAN) és a trigeminális idegből származó aceton (ACE) szaglását is. A VAN jeleit kizárólag a szagló nyálkahártya közvetíti, míg az ACE jeleit a légzőhám is továbbítja [Cain és Murphy, 1980 ; Hummel és Welge‐Luessen, 2006 ; Savic et al., 2002 ].
Spoiler: ha nem akarod végigolvasni a lenti kutatás hosszú tudományos részleteit, akkor az eredmény az lett, hogy az egyes feromonok érzékelése a szagló hagymán (MOB) keresztül történik az emberekben és nem (vagy nem csak) a Vemonorosal organ (VNO) szerven keresztül.
Tehát a kutatás bebizonyította, hogy az ember érzékeli a feromonokat a szaglóhagymán keresztül.
Így zajlott az emberi feromon érzékelése kutatás
A vizsgálatba tizenkét jobbkezes, nem dohányzó, heteroszexuális férfit (21–42 évesek) toboroztak, akiknél orrpolip okozta krónikus anozmiát diagnosztizáltak.
A polipózist fül-orr-gégész szakorvos diagnosztizálta, és endoszkóppal vizualizálták, amely a szaglóhám apikális elzáródását mutatta, de a kaudális irányban elhelyezkedő vomeronazális gödrök épek maradtak. A polipózison kívül az alanyok jó egészségi állapotban voltak, és a részletes kórtörténetük alapján nem számoltak be neuropszichiátriai rendellenességekre vagy anozmiára vonatkozó öröklődő tényezőről.
Az anozmia átlagos időtartama 4,7 év volt (tartomány: 1,5–10 év). A szaglási diszfunkciót polipózis okozta, és a kórtörténetben nem szerepelt trauma, felső légúti fertőzés vagy öröklődés okozta hipo- vagy anozmia. A résztvevők egyikénél sem volt veleszületett anozmia, sem specifikus szagokra vonatkozó anozmia a kórtörténetben. A szaglási küszöböket n- butil-alkohol teszttel mérték a korábban leírtak szerint [Savic et al., 2000 ], és fenil-etil-alkohol teszttel igazolták [Bensafi et al., 2007 ]. A vizsgálatba csak azokat a betegeket vonták be, akik hígítás nélkül sem érzékelték a két szag egyikét sem.
A trigeminális ideg érzékenységét is felmértük a vizsgálati alanyok azon képessége alapján, hogy képesek-e lateralizálni a bal vagy a jobb orrlyukba irányuló ingereket. Erre a célra 99%-os eukaliptolt juttattunk az egyik orrlyukba egy 10 ml-rel töltött üvegedényben, amelynek kiöntőjét az egyik orrlyukba helyeztük (Sigma Chemical, Deishofen, Németország), [Hummel et al., 2003 ]. Szagtalan levegőt juttattunk párhuzamosan, és egy külön üvegen keresztül, amelynek kiöntőjét a másik orrlyukba helyeztük. Az utasítás az volt, hogy jelezzék az érzékelt irritáció, szag vagy égő/viszkető érzés oldalát, amely az eukaliptol beadásának oldalához kapcsolódott. Összesen 10 ingert alkalmaztunk bekötött szemű alanyokon, körülbelül 60 másodperces ingerek közötti intervallummal. A jobb és bal orrlyuk ingerlését ellensúlyoztuk és randomizáltuk. Minden inger után megkérték a vizsgálati alanyt, hogy azonosítsa azt az orrlyukat, ahol a szaglóanyagot észlelte. Az elemzésben a helyes azonosítások összegét használtuk.
A kontrollcsoport 12 jobbkezes, nem dohányzó, heteroszexuális, életkor és iskolai végzettség szerint illeszkedő férfiból (21–36 év) állt, akiknél semmilyen felső légúti probléma, neuropszichiátriai rendellenesség vagy öröklődés nem állt fenn. Közülük 11 adatot egy korábbi tanulmányunkban [Berglund et al., 2008 ] mutattunk be. A tanulmányt a helyi etikai és biztonsági sugárzási bizottságok jóváhagyták.
Szagok
A Sigma Chemical (Deishofen, Németország) által gyártott tiszta (99%) vanillint (4-hidroxi-3-metoxi-benzaldehid) és acetont (C3H6O, HPLC 99,9%) folyékony formában, hígítás nélkül használtunk. A PET-vizsgálatok során végzett korábbi tanulmányainkhoz hasonlóan az AND és az EST kristályos formában és szagos (200 mg, Steraloids, Newport, USA) formában volt jelen [Savic et al., 2001 ].
Feltételek és kísérleti eljárás
A PET-vizsgálatokat három különálló körülmény között végezték: birhinális, szagtalan levegő passzív szaglása (AIR), aceton (ACE), vanillin (VAN), AND és EST. Acetont és vanillint használtak annak vizsgálatára, hogy az anosmikusok hogyan dolgozzák fel a tiszta szagló- és a kevert szagló- + trigeminális szaglóanyagokat [Savic et al., 2002 ]. Ezeket a szaglóanyagokat annak megállapítására használták, hogy az anosmikusok szaglóanyagokból származó jelek átvitelének képtelensége a szaglónyálkahártyára korlátozódik, és nem érinti a légzőszervi orrhámot. Az AND egy szteroid, mint az EST, és azért vizsgálták, hogy az anosmikus férfiaknál az EST általi lehetséges aktiváció az adott vegyülethez, és nem annak szteroid szerkezetéhez kapcsolódik-e.
Az alanyonkénti vizsgálatok száma 14-re korlátozódott. Minden alany szagtalan levegőt, AND-t és EST-t érzett három vizsgálat során; az alanyok 50%-a három vizsgálat során VAN-t, kettőben pedig ACE-t érzett; a fennmaradó alanyoknál háromszor ACE-t, kétszer pedig VAN-t mutattak be. A feltételek sorrendje kiegyensúlyozott volt az alanyok között. A vizsgálatokat mozgási műtermékek miatt kizártuk a további elemzésből. A vizsgálatokat mozgási műtermékek miatt kizártuk a további elemzésből.
A szagosító anyagokat üvegpalackokban és vattapamacsban tartották. Amikor a vizsgálat levegőszagolásról szólt, a pálcát desztillált vízzel áztatták. Minden egyes tárgyat külön vizsgálat során mutattak be, 10 mm távolságra az orrlyukaktól. A szagadaptáció minimalizálása érdekében az adott tárgyat a vizsgálat során 15 másodpercig, összesen négyszer mutatták be. Mindegyik bemutatást 5 másodperces szünet követte, amikor a vizsgálati helyiségben lélegeztették a levegőt, ahol a vizsgálati állvány közelében elhelyezett szívófülke biztosította a folyamatos szívást és levegőfrissítést. Az első bemutatás 5 másodperccel a 15O —H2O bólus injekciója után kezdődött . Egy stimulációs ülés 80 másodpercig, minden PET-mérés 60 másodpercig tartott.
A körülményektől függetlenül minden egyes prezentációt a vizsgálati alany jobb mutatóujjának megérintésével jeleztek. Az alanyokat tájékoztatták, hogy vagy szagot, vagy szagtalan levegőt fognak érezni, anélkül, hogy ismerték volna a tárgyak típusát vagy sorrendjét. Utasítást kaptak, hogy passzívan lélegezzenek (ne szippantsanak, még akkor sem, ha nem érzékelnek szagot), és a bemutatott tárgy érzékelésére koncentráljanak, annak jellemzőinek elemzése nélkül. Számos előkészítő pszichofizikai kísérlet során az alanyokat más szagokkal képezték ki a szkennerben, hogy megismerkedjenek egy kísérleti eljárással. Minden szkennelés során légzésmozgásokat rögzítettek az alsó mellkas körül elhelyezett nyúlásmérő bélyeggel (Comair AB, Stockholm). A légzési frekvenciát (lélegzetvétel/perc) és amplitúdót folyamatosan rögzítették a VAN, ACE, AND, EST és AIR minden egyes prezentációja előtt és alatt. A milliméterben mért amplitúdó közvetlenül összefügg a nyúlásmérő szalag nyújtásának mértékével, azaz az alsó mellkas kerületének növekedésével.
A PET-adatok elemzése
A PET-kísérleteket minden alanynál mágneses rezonancia képalkotás (MRI) előzte meg (1,5 Tesla GE szkenner; 3D SPGR; TE = 5 ms, TR = 21 ms; FOV = 256 mm). A használt PET-szkenner a CTI‐Siemens ECAT EXACT HR szkenner volt, 3D módban. A PET-vizsgálatok során minden alany körülbelül 12 mCi 15O — H2O – t tartalmazó bólus injekciókat kapott . A PET-vizsgálatokat rögzített fejjel [Bergstrom et al., 1981 ], bedugott fülekkel és becsukott szemekkel végezték. A PET-szobában a hőmérsékletet és a légnyomást minden kísérlet során standardizálták (23°C, 997 hPa), a szagok okozta szennyeződést pedig a szkennerhez csatlakoztatott szívóberendezéssel küszöbölték ki. A betegeket és a kontrollcsoportot átfedő időszakban, a nap azonos szakában, azonos körülmények között, ugyanazon kísérletezők vizsgálták.
A relatív regionális agyi véráramlást (rCBF) a felvétel megkezdése után 60 másodpercben számítottuk ki 10 mm-es Gauss-szűrő segítségével. A PET-képeket előfeldolgozásnak vetettük alá, beleértve az anatómiai standardizálást és a globális normalizálást 50 ml/100 g/perc értékre, az SPM2 statisztikai csomag (Wellcome Foundation, London) segítségével [Frackowiak, 2004 ; Friston et al., 1999 ; Savic et al., 2000 , 2001 ].
A szignifikáns aktivációkat először minden külön csoportban egycsoportos véletlenszerű hatásanalízissel értékeltük (T-küszöb 0,001, korrigált P -érték <0,05) a következő kontrasztok esetén: AND–AIR és vv; EST–AIR és vv; VAN–AIR és vv; ACE–AIR és vv. Ezután a csoportkülönbségeket kétcsoportos véletlenszerű hatásanalízissel teszteltük. A szignifikáns klasztereket a magassági küszöbértéknél , T = 0,01-nél, korrigált P < 0,05-nél (SPM2, Wellcome Foundation, London) számítottuk ki, hogy elkerüljük annak kockázatát, hogy a kisebb csoportkülönbségek észrevétlenek maradjanak.
A csoportok közötti értékelések korábbi eredményeinkre támaszkodtak, amelyek azt mutatták, hogy az AND- és EST-függő aktivációk a hipotalamuszra és a szaglóagyra korlátozódtak. Tekintettel arra, hogy az SPM statisztikák meglehetősen konzervatívak, és a II. típusú hibák elkerülése érdekében egy téglalap alakú maszkot használtunk, amely csak az agy vízszintes szakaszait fedte le Z = +20 és Z = −20 között. Az ACE-t és VAN-t tartalmazó kontrasztok (ACE-AIR, VAN-AIR és vv) esetében a keresési tér az egész agy volt.
Eredmények elemzése
Az érdeklődésre számot tartó térfogat (VOI) analízist alkalmazták annak értékelésére, hogy az anozmiás férfiak hasonló módon aktiválták-e az elülső hipotalamuszt, mint a kontrollcsoport tagjai. Mivel ennek a régiónak az anatómiai határait nehéz MRI-vel meghatározni, úgynevezett funkcionális VOI-t alkalmaztak. Ezt a VOI-t egy korábbi, heteroszexuális férfiakon végzett vizsgálatban generálták, akiket hasonló kísérleti eljárással vizsgáltak [Savic et al., 2001 ]. A VOI-t a hipotalamusz klaszter alkotta, amelyet az EST-AIR kontrasztanyag generált. A PET-képekből definiálták, amelyeket ugyanarra a standard agyra formáztak újra, mint amilyeneket ebben a vizsgálatban is használtak, így közvetlenül átvihető volt az egyes PET-képekre. Az rCBF-et minden alanynál először normalizálták az 50 ml/perc/100 g globális agyi átlaghoz, és az ebben a VOI-ban található átlagos rCBF-et minden alany összes vizsgálatára kivonták [Savic et al., 2001 ]. Az egyes körülmények között extrahált átlagos rCBF lehetséges különbségeit ezután összehasonlítottuk az EST és AIR, AND és AIR, VAN és AIR, valamint ACE és AIR között minden alanycsoport esetében párosított t- próbákkal. A szabadsági fokok száma (df) 11 volt. A P- szint 0,05 volt, azon a hipotézisen alapulva, hogy csak az EST-AIR eredményez szignifikáns aktivációt. Az anosmikusok és a kontrollok közötti lehetséges különbségeket ezen aktivációk tekintetében ismételt méréses ANOVA-val értékeltük, az alanycsoportot közötti, a szaglóanyag típusát pedig belső faktorként használva. Amikor szignifikáns interakciót észleltünk, az eredményeket kontrasztokkal tovább vizsgáltuk annak meghatározására, hogy melyik szaglóanyag típusa határozta meg a megfigyelt interakciót. A P- értékeket <0,05 esetén tekintettük szignifikánsnak.
A pszichofizikai adatok elemzése
Az adott vizsgálat során a légzési frekvenciát és amplitúdót az egyes vizsgálatok előtti 2 perc nyugalmi állapothoz viszonyítottuk (alapértéknek tekintettük). Minden vizsgálathoz kiszámítottuk a légzési frekvencia és a légzési amplitúdó szorzatát, és ezt az indexet használtuk a légzési térfogat alapértékhez viszonyított változásának leírására. Az ismételt méréses ANOVA-ban a csoportokat a változó és a négy szagos vegyület közé, a levegőt pedig a változókon belüli változóként soroltuk be.
EREDMÉNYEK
A kontrollcsoportok szagküszöbértékei normálisak voltak (5,0 × 10⁻⁶ ± 2,0 × 10⁻⁶ M ). Az aceton helyesen lateralizált oldalán a kontrollcsoportban 9,3 ± 0,4, az anosmikusokban (ns) pedig 8,7 ± 0,8 volt.
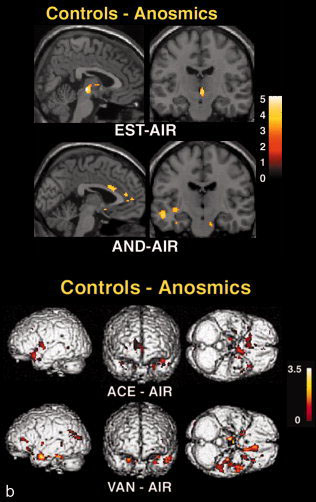
Csoportspecifikus aktivációk illusztrációja a feltételezett feromonokkal. ( Felső ) Agyi aktiváció AND és EST szaglása során. Az aktivált régiók klaszterei a standard MRI agy ( spm99 ) középszagittális síkján helyezkednek el. Az EST klaszter alsó része homoszexuális férfiaknál az amygdalában és a piriform kéregben található. ( Alsó ) A csoportok közötti szignifikáns különbségek. Az ábrán a kétcsoportos véletlenszerű hatásanalízissel számított klaszterek láthatók. A Sokoloff színskála az aktiváció mértékét tükröző z értékeket szemlélteti. Csak a szignifikáns aktivációk láthatók. Mivel ugyanazt az agyterületet választottuk, az ábrák nem mindig szemléltetik az egyes állapotok maximális aktivációját.
Aktiválások
Amint azt korábban beszámoltuk, a heteroszexuális férfi kontrollcsoportok az elülső hipotalamuszban, a bal oldali amygdalában és a piriform kéregben csoportosulásokat mutattak EST szaglásakor ( I. táblázat ) [Savic et al., 2001 ]. Az anozmikusok ezzel szemben nem mutattak agyi aktivációt.
Aktiválások és deaktiválások anosmikai és kontrollrendszerekben
| Régió | Vezérlők | Anosmikai | ||||
|---|---|---|---|---|---|---|
| Z szint | Méret, cm 3 | Koordináták | Z szint | Méret, cm 3 | Koordináták | |
| Aktivációk anosmikai és kontroll folyamatokban | ||||||
| ACE-AIR | ||||||
| R insuláris kéreg (tartalmazza az amygdiát + a pirif kérget + a farokmagot + az elülső cingulációt) | 6.5 | 9.0 | 38, −10, 6 | |||
| R farokcsont | ||||||
| L-szigetes kéreg (beleértve a posztcentrális gyrust) | 5.2 | 5.0 | −30, −6, 0 | 3.0 | 1.4 | −50, 10, 30 óra |
| Hangya cinguláris | 3.5 | 5.6 | −22, 44, 8 | |||
| Hipotalamusz és talamusz b | 3.5 | 2.8 | 16, −10, −20 | 4.2 | 2.2 | 2, −36, −6 |
| 4.2 | 5.0 | 8, −32, −20 | 12, 2, 16 | |||
| 5.1 | 2.4 | −2, −2, −6 | ||||
| Kisagy | 4.3 | 0,8 | −48, −50, −28 | |||
| VAN-AIR | ||||||
| R pirif + amigd | 5.1 | 1.6 | 20, −4, −20 | |||
| L pirif + amigd | 4.8 | 0,9 | −20, −4, −16 | |||
| L orsó alakú gyrus | 4.3 | 1.3 | −48, −42, −26 | |||
| ÉS-LEVEGŐ | ||||||
| R pirif + amigd | 4.5 | 0,9 | 36, −14, −6 | |||
| R lingularis + fusiform gyrus | 4.2 | 1.2 | 18, −60, −24 | |||
| EST-AIR | ||||||
| Elülső hipotalamusz | 4.6 | 1.2 | 6, −12, 2 | |||
| L pirif + amigd | 4.5 | 1.0 | −22, −6, 24 | |||
| Deaktivációk anosmikában és kontrollokban | ||||||
| AIR-ACE | ||||||
| R felső temporális gyrus | 4.5 | 1.6 | 54, −12, −6 | 4.5 | 1.6 | 54, −12, −6 |
| Jobb, bal középső temporális gyrus | 4.4 | 1.2 | 38, −42, −14 | 4.5 | 3.2 | 42, −68, 8 |
| 3.3 | 3.2 | 38, −18, −20 | 38, −62, 12 | |||
| Parieto-occipitális kéreg | 4.5 | 33 | 12, −68, 16 | 5.0 | 6.2 | −10, −82, 34 |
| 3.3 | 2.8 | −38, −36, 30 | ||||
| LEVEGŐ-ÉS | ||||||
| Mediális prefrontális kéreg | 3.7 | 2.5 | 4, 46, 20 | |||
| AIR-EST | ||||||
| Elülső cinguláris | 3.6 | 3.3 | 6, 58, 0 | |||
| LÉGI VAN | ||||||
| Mediális prefrontális kéreg | 4.5 | 5.6 | 38, 46, 8 | |||
| 4.8 | 3.6 | −14, 45, 2 | ||||
| Középső temporális gyrus | 4.4 | 1.2 | −40, −44, 6 | |||
| Cuneus, orsó alakú gyrus | 4.3 | 3.6 | 46, −74, −16 | |||
Egycsoportos véletlenszerű hatásanalízissel számított értékek, P = 0,001 magassági küszöbértékkel és P < 0,05 korrigálva. A Talairach-koordináták a csúcsaktivációt jelzik; a jelzett régiók az adott klaszter lefedettségét írják le. pirif, piriform kéreg; amygd, amygdala.
A kontrollcsoportban az AND és a VAN szaglása az amygdala piriform, az agranuláris insuláris kéreg és a fusiform gyrus egyes részeinek jelentős aktivációjával járt. Az acetonnak való kitettség további klasztereket eredményezett az elülső cingulárisban, az agytörzsben (beleértve a trigeminális magot is), a talamuszban (a ventromediális mag és a pulvináris), a szenzorimotoros kéregben (az arc topográfiai reprezentációjának felel meg) és a kisagyban. Az anoszmikusok nem mutattak aktivációt VAN vagy AND esetén, és jelentős klaszterek csak az ACE-AIR esetén jelentek meg. Ezeket az elülső cingulárisban, a talamuszban (a ventromediális mag és a pulvináris), az agytörzsben (a trigeminális magot lefedve) és a szenzorimotoros kéregben (az arc topográfiai reprezentációja) észlelték. Figyelemre méltó, és a kontrollcsoporttól eltérően, nem észleltek amygdala, piriform vagy insuláris klasztereket ( I. táblázat ). Kisagyi aktiváció sem volt megfigyelhető.
Deaktiválások
A deaktivációkat a normalizált rCBF csökkenéseként definiáltuk szaglóanyagok szaglása során a levegőhöz képest. ACE-vel kapcsolatos deaktivációkat észleltünk a kontrollokban, valamint az anosmikusokban a temporális és a parieto-occipitális kéregben. A kontrollok további deaktivációkat mutattak a frontopoláris és a dorsolaterális prefrontális kéregben. Az EST, az AND és a VAN szaglása során a deaktivációkat csak a kontrollokban észleltük; ezek a frontopoláris kéregben és a dorsolaterális prefrontális kéreg egyes részein helyezkedtek el ( I. táblázat ).
Csoportos összehasonlítások
A várakozásoknak megfelelően a kontrollok és az anosmikusok (kontrollok–anosmikusok) közötti kontraszt szignifikáns klasztert mutatott az elülső hipotalamuszban az EST–AIR esetében, valamint a bal oldali amygdalában is ( II. táblázat, 1. ábra a). Az AND–AIR, VAN–AIR és ACE–AIR megfelelő csoport-összehasonlítása csak az amygdalában, a priformban és az elülső insularis kéregben mutatott klasztereket. A VAN–AIR és ACE–AIR tekintetében végzett csoport-összehasonlítások további klasztereket mutattak az elülső cingulárisban és az elülső talamuszban, valamint az ACE‐AIR szenzorimotoros kéregében is ( II. táblázat, 1. ábra b). Az anosmikusok és a kontrollok (anosmikusok–kontrollok) összehasonlításakor szignifikáns klaszterek csak az ACE–AIR esetében jelentek meg; ezek a prefrontális kéregben helyezkedtek el, amely az orbitofrontális kéreg egyes részeit fedi le, és a parietális kéregben: ez utóbbi régió átfedésben volt a kontrollokban az ACE-vel kapcsolatos deaktivációval ( I. és II. táblázat ).
| Régió | Kontrollok – anosmikák | Anosmika – kontrollok | ||||
|---|---|---|---|---|---|---|
| Z szint | Méret, cm 3 | Koordináták | Z szint | Méret, cm 3 | Koordináták | |
| ACE-AIR | ||||||
| R amygd + pirif kéreg | 4.6 | 3.9 | 20, 4, −14 | |||
| L-szigetes kéreg + amigda | 3.6 | 0,7 | −36, −24, 10 | |||
| 3.4 | 1.2 | −12, 6, −10 | ||||
| Hangya cinguláris kéreg | 3.9 | 1.3 | −4, 42, 0 | 4.5 | 1.7 | −18, 42, −4 |
| 12, 62, 14 | ||||||
| Subcallosum | 3.0 | 0,4 | 18, 20, −18 | |||
| Precentrális + posztcentrális gyrus | 2.9 | 0,4 | −54, 0, 8 | 4.0 | 1.8 | −40, −6, 52 |
| Alsó frontális gyrus | 3.8 | 2.4 | −44, 20, 10 | |||
| Cuneus | 3.0 | 0,8 | −4, −84, 24 | |||
| VAN-AIR | ||||||
| L amygda + pirif + agranuláris insuláris kéreg + elülső talamusz | 3.9 | 5.3 | −44, 0, −26 | |||
| Cingularis kéreg | 3.8 | 1.7 | −6, 30, −8 | |||
| ÉS-LEVEGŐ | ||||||
| R amygd + pirif kéreg | 4.6 | 3.9 | 20, 4, −14 | |||
| R lingularis + fusiform gyrus | ||||||
| EST-AIR | ||||||
| Elülső hipotalamusz | 4.0 | 0,5 | 2, −15, −4 | |||
| L amygda + pirif kéreg | 4.5 | 1.0 | −22, −6, 24 | |||
Két csoportos véletlenszerű hatásanalízissel számított értékek, P = 0,01 magassági küszöbértékkel , korrigált P < 0,05 értékkel az ACE-AIR és VAN-AIR esetében, valamint korrigálatlan P < 0,05 értékkel az AND-AIR és EST-AIR esetében. A Talairach-koordináták a csúcsaktivációt jelzik; a jelzett régiók az adott klaszter lefedettségét írják le. J, jobb; L, bal; amigda, amigdala; pirif, piriform kéreg.
A VOI-analízis szignifikáns csoportkülönbséget mutatott ( P = 0,0002, df = 3, teljesítmény = 0,988, F = 16,3). Ezt az EST-AIR tette ki, amely szignifikánsan magasabb aktivációt mutatott a kontrollokban ( P < 0,0001).
Légzési válaszok
A számított légzési indexben nem volt szignifikáns csoportkülönbség ( F = 0,13; P = 0,7: df = 1) vagy csoport × vegyület interakció ( F = 1,38; P = 0,2; df = 4). Lásd az S1. ábrát a kiegészítő információkban.
Következtetések
A férfi kontrollcsoporttal ellentétben, akiknél az elülső hipotalamuszban egyértelmű EST-aktivációt figyeltek meg, az anosmiás férfiaknál nem észleltek EST-aktivációt. Ezt a megállapítást SPM-analízissel támasztották alá, és a VOI-analízissel megerősítették. Az alkalmazott módszereket számos korábbi tanulmányunkban ismertettük, amelyek konzisztens és reprodukálható eredményeket hoztak [Berglund et al., 2006 ; Savic et al., 2001 , 2002 , 2005 ]. Ezért itt a továbbiakban nem részletezzük őket.
Az orrpolipózis alapján végzett anozmia vizsgálatunk oka az volt, hogy a polipok csak a szaglónyálkahártyát érintik. A környező hámréteg kisebb mértékű érintettsége nem zárható ki, de valószínűtlennek tűnik, tekintve, hogy minden anozmiás beteg acetont érzett, és egyértelmű agyi aktivációt mutatott ezzel a vegyülettel. Továbbá a VNO az orrfenéktől távol helyezkedik el, amely az egyetlen olyan terület, amelyet általában a polipok érintenek, és az endoszkópos vizsgálatok nem utaltak a légzőszervi nyálkahártya rendellenességére. Ezenkívül egyik betegünk sem számolt be rhinitisről, a rendelkezésre álló szakirodalom szerint a polipózis elsősorban krónikus nasorhinitissel összefüggésben érinti az orrnyálkahártyát [Slater et al., 1996 ]. A betegek és a kontrollcsoport életkora hasonló volt, és nem járulhatott hozzá a szagérzékenységbeli különbségekhez. Az anozmiának sem lehetett más etiológiája a polipózison kívül, tekintve, hogy egyik beteg sem öröklött általános vagy szelektív anozmiára, és hogy minden beteg normális szaglásról számolt be a pangás előtt.
A várakozásoknak megfelelően az anosmikók abban különböztek a kontrollcsoporttól, hogy nem aktiválták a VAN, AND, EST, valamint az ACE vonatkozásában a szaglási feldolgozáshoz kapcsolódó régiókat. Így a szaglóanyag típusától vagy az inger kémiai szerkezetének típusától függetlenül nem észleltek szignifikáns klasztereket a szaglóhálózatokban. Épp ellenkezőleg, a betegek aktiválták a szaglóanyagok trigeminális idegi feldolgozásában részt vevő neuronális áramköröket [Boyle et al., 2007 ; Savic, 2002 ], megerősítve, hogy a szaglószerv, de nem a légzőhám érintettségében szenvedtek. Az ACE-vel történő aktivációk valamivel kevésbé voltak kifejezettek az anosmikóknál a kontrollcsoporthoz képest ( II. táblázat, 1b . ábra ). Ez a különbség nem tulajdonítható a légzési mintázatnak, amely nem különbözött szignifikánsan a két csoport között. Az egyik lehetőség az, hogy az anosmikók csökkent érzékenységet mutattak az intranazális trigeminális ingerekre az aceton trigeminális komponensének szaglási fokozódásának elvesztése miatt [Cain és Murphy, 1980 ; Cashion et al., 2006 ; Gudziol és munkatársai, 2001 ; Hummel és munkatársai, 1996 ]. Egy másik elmélet szerint az anosmikusokban a trigeminális idegrendszer feldolgozása funkcionálisan átszerveződött központi szinten, amint azt az anosmikusokban a gáz halmazállapotú CO2 – ami egy szagtalan trigeminális inger – jelenlétében csökkent agyi aktivációról szóló beszámolók is mutatják [ Iannilli és munkatársai, 2007 ]. A két mechanizmus nem zárja ki egymást, és együttesen is létezhet.
Elméletileg a heteroszexuális férfi kontrollcsoportunk feldolgozhatta az EST-ből származó jeleket VNO-n, a szaglónyálkahártyán vagy a vénás véren keresztül (miután az EST felszívódott az orr érrendszerébe). A humorális transzport valószínűtlennek tűnik, mivel sokkal lassabb, mint a jelenleg megfigyelt agyi aktivációk. Például a bore feromon androsztenol orron keresztüli alkalmazása után a maximális plazmakoncentráció 40 és 50 perc között figyelhető meg az alkalmazás után, míg a mi kísérleteinkben az agyi aktivációt már 30 másodpercen belül kimutatták [Stefanczyk‐Krzymowska et al., 2000 ]. Továbbá, ha az EST-jelek humorális úton jutnának el a hipotalamuszba, az aktivációs mintázat hasonló lenne a kontrollcsoportban és az anozmikusokban, ami nem így történt.
A jelenlegi eredmények azt a hipotézist is elvetik, hogy az EST feromon komponenséből származó jeleket a VNO közvetíti; inkább azt a nézetet erősítik, hogy egészséges férfiaknál az ebből a komponensből származó jeleket a MOB feldolgozhatja a szaglóhámján keresztül. Érdekes módon számos bizonyíték utal arra, hogy a MOB szerepet játszhat a feromonjelek detektálásában állatokban is. A MOB-ot a görényekben [Woodley és Baum, 2003 ] és a hörcsögökben [O’Connell és Meredith, 1984 ] feromonok aktiválják. A VNO ablációja nincs hatással a nyulak szopási viselkedésére [Hudson és Distel, 1986 ] vagy a hím hörcsögök párzási viselkedésére [Pfeiffer és Johnston, 1994 ]. A műtéti úton eltávolított VNO-val rendelkező hím egerek továbbra is képesek megkülönböztetni a vizelet szagát a hímek és az ivarzás alatt álló nőstények között. Hasonlóképpen, a VNO eltávolítása a nőstény egerekből nem rontja a hímek és nőstények megkülönböztetésének képességét, míg a szaglóhám ZnSO4-gyel történő elpusztítása csökkenti a szexuális viselkedésüket [Keller et al., 2006 ]. A MOB-ot feromonok aktiválják görényekben és hörcsögökben is [O’Connell és Meredith, 1984 ; Woodley és Baum, 2004 ].
A feromonérzékelő receptorok egy speciális családját, az úgynevezett „nyomaminosszociált receptorokat” (TAAR) nemrégiben észlelték az egér szaglóhámjában. Érdekes módon az ezeket a receptorokat kódoló gének emberekben is jelen vannak [Liberles és Buck, 2006 ], ami összhangban van Rodrigez és munkatársai [ 2000 ] korábbi jelentésével egy funkcionális feromonreceptor gén expressziójáról az emberi szaglónyálkahártyában. Ezek a, mégis anekdotikus eredmények együttesen tovább alátámasztják azt a hipotézist, hogy az emberek, számos más emlőshöz hasonlóan, képesek a feromonjelek feldolgozására a szaglónyálkahártyán keresztül. Javasoljuk további vizsgálatokat ezeken a területeken.
Köszönetnyilvánítás
Köszönettel tartozunk Dr. Gulyás Balázsnak a PET-jelzőanyag beadásáért a kísérletek része során.
A természetes feromon hatás – vonzerő növelése
Férfiak és nők esetében is a súlyzós edzés, nagy mértékű fizikai munka végzése során beindul a feromon termelődés. Izzadás során a bőrmirigyek feromont termelhetnek. Az így termelt feromon, sajnos nem marad velünk hosszú távon hiszen a tisztálkodás során jelentős mennyiséget lemosunk magunkról, ezért nagy mértékben elveszik a saját testünk által termelt feromon.
Tévhit, hogy a cink fogyasztása segíti a feromon termelést.
Talán a legegyszerűbb megoldás a természetes feromonjaid felerősítésére a palackba zárt extra csáberő: használj eredeti feromon parfümöt, ez az egyik legbiztosabb módja, hogy biztosítsd az extra feromon jelenlétét magad körül a mindennapjaid során. A parfüm pontosan tartalmazza azt a fajta feromont, amire szükséged van, mindemellett egész nap tartó illatot biztosít.
Tudományos feromon kísérletek és hivatalos orvosi publikációk
A kutatásban és a cikkben felhasznált tudományos irodalom jegyzéke
[1] A kemoszenzoros neuronok fejlődésének és regenerációjának összehasonlító immuncitokémiai vizsgálata vomeronasalis rendszerben, Neurobiológiai és anatómiai részleg, Niigata, Japán – Trotier D, Eloit C. „The vomeronasal system of primates: a comparative approach.” Chem Senses. 2011;36(1):1-15. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[2] Anatómiai bizonyítékok a vomeronasalis szerv endokrin aktivitására emberekben – Stensaas LJ et al. „Anatomical evidence for the existence of the vomeronasal organ in adult humans.” J Comp Neurol. 1991;311(3):301-13. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[3] A nemi hormonszerű vegyületek szaga nemileg differenciált hipotalamusz aktivációt okoz az emberben, Emberi Agykutatási Osztály, Idegtudományi Osztály, Karolinska Institute, Stockholm, Svédország – Savic I et al. „Smelling of odorous sex hormone-like compounds causes sex-differentiated hypothalamic activations in humans.” Neuron. 2001;31(4):661-8. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[4] Az emberi vomeronasális szerv működése: a legjobb és legrosszabb esetek kritikus áttekintése, Biológiai Tudományok Tanszék, Florida Állami Egyetem – Meredith M. „Human Vomeronasal Organ function: A critical review of best and worst cases.” Chem Senses. 2001;26(4):433-45. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[5] Ultraszenzitív feromon kimutatás emlős vomeronasalis neuronokkalAnatómiai és Neurobiológiai Tanszék, University of Maryland School of Medicine, USA – Leinders-Zufall T. et al. (2000). Ultrasensitive pheromone detection by mammalian vomeronasal neurons. Nature, 405(6788), 792–796. [PubMed: 10866200] Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[6] Megváltozott szexuális és szociális viselkedés, Biokémiai és molekuláris biofizikai tanszék, College of Physicians and Surgeons, Columbia Egyetem, New York, USA – Leypold B.G. et al. (2002). Altered sexual and social behaviours in trp2 mutant mice. Proceedings of the National Academy of Sciences, 99(9), 6376–6381. [PubMed: 11972034] Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[7] MHC I. osztályú peptidek, mint kemoszenzoros jelek a vomeronasalis szervben, Anatómiai és Neurobiológiai Tanszék, University of Maryland School of Medicine, Baltimore, USA – Leinders-Zufall T. et al. (2004). MHC class I peptides as chemosensory signals in the vomeronasal organ. Science, 306(5698), 1033–1037. [PubMed: 15528444] Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[8] Az emberi kemojelek vizsgálata, Neurobiológiai Osztály, Weizmann Tudományos Intézet, Rehovot, Izrael. – Sobel N. et al. (1999). Human chemosignals: integrating neuroimaging and psychophysiology. Chemical Senses, 24(4), 447–456. [PubMed: 10479803] Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[9] Szex és az orr: emberi feromon válaszok, Fül-orr-gégészeti osztály, Kettering General Kórház, Rothwell Road, Kettering, Northants, UK – Bhutta M.F. (2004). Sex and the nose: human pheromonal responses. Journal of the Royal Society of Medicine, 97(6), 268–274. [PubMed: 15173330] Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[10] A feromonoktól a viselkedésig, Idegtudományi Tanszék, Parmai Egyetem, Trieste, Olaszország – Tirindelli R et al. “From pheromones to behavior.” Physiol Rev. 2009;89(3):921–956. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[11] Feromon jelek molekuláris kimutatása: a génektől a viselkedésig, Molekuláris és sejtbiológiai osztály, Harvard Egyetem, Cambridge, USA – Dulac C, Torello AT. “Molecular detection of pheromone signals in mammals: from genes to behaviour.” Nat Rev Neurosci. 2003;4(7):551–562. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[12] Az emberi feromonok valószínűsége, Nature folyóiratban megjelent tanulmány – Comfort A. Likelihood of human pheromones. Nature 1971; 230: 432–3. PMID: 4932036 DOI: 10.1038/230432a0 Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[13] A feromonok emberre gyakorolt hatásának neurobiológiai korrelációi – Savic I et al. “Neurobiological correlates of pheromonal influence on humans.” J Comp Neurol. 2005;493(3):457–466. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[14] A párválasztás és a romantikus szerelem idegi mechanizmusai – Fisher HE et al. “The neural mechanisms of mate selection and romantic love.” J Comp Neurol. 2006;493(1):58–62. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[15] A szerelem neurobiológiája, University College, Anatómia Tanszék, London, Egyesült Királyság, Zeki S. “The neurobiology of love.” FEBS Lett. 2007;581(14):2575–2579. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[16] MHC-függő párpreferenciák emberekben, Abteilung Verhaltensökologie, Universität Bern, Svájc – Wedekind C et al. “Evidence for MHC-correlated perfume preferences in humans.” Proc Biol Sci. 1995;260(1359):245–249. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[17] Az androstenol – egy szteroid eredetű szag aktiválja a hipotalamusz a nőknél. – Savic I, Berglund H. “Androstenol–a steroid derived odour activates the hypothalamus in women.” J Neurosci. 2001. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[18] Az androsztadienon által kiváltott agyi aktiváció heteroszexuális nőknél. – Gulyas B et al. “Cerebral activation induced by androstadienone in heterosexual women.” NeuroReport. 2004. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[20] A nem kemoszenzoros kommunikációja két emberi szteroidon keresztül, szexuálisan dimorf módon, Kínai Tudományos Akadémia Pszichológiai Intézete, Peking – Zhou W et al. “Chemosensory communication of gender through two human steroids in a sexually dimorphic manner.” Curr Biol. 2014. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[21] Feromon jelátvitel emberekben – Kutatók: Dr. Gulyás Balázs, Ivanka Savic, Ebba Hedén‐Blomqvist, Hans Berglund – Stockholmi Agykutató Intézet, Klinikai Idegtudományi Osztály, Svédország PMCID: PMC6870699 Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[22] Kísérlet a szagérzékelés nemi különbségeire vonatkozóan – Pszichológiai laboratórium, Utrecht Egyetem, Hollandia // Koelega, H. S., a Koster, E. P. (1974). Some experiments on sex differences in odor perception. Ann N Y Acad Sci, 237(0), 234-246. The New York Academy of Sciences
[23] Egy emberi szexuálszteroid eredetű vegyület szaglása dózisfüggő módon befolyásolja a hangulatot és az autonóm arousalt. Bensafi M, Tsutsui T, Khan R, Levenson RW, Sobel N (2004): Psychoneuroendocrinology 29: 1290–1299. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[24] A szagok idegi reprezentációját a szagkódolás során a trigeminális inger jelenléte modulálja. Bensafi M, Frasnelli J, Reden J, Hummel T (2007): Clin Neurophysiol 118: 696–701. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[25] Agyi válasz feltételezett feromonokra leszbikus nőknél. Berglund H, Lindstrom P, Savic I (2006): Proc Natl Acad Sci USA 103: 8269–8274. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[26] Férfiból nővé alakuló transzszexuálisok nemi atipikus hipotalamusz aktivációt mutatnak szagos szteroidok szaglásakor. Berglund H, Lindstrom P, Dhejne-Helmy C, Savic I (2008): Cereb Cortex 18: 1900–1908. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[27] Fejrögzítő eszköz reprodukálható pozícióillesztéshez transzmissziós CT-ben és pozitronemissziós tomográfiában. Bergstrom M, Boethius J, Eriksson L, Greitz T, Ribbe T, Widen L (1981): J Comput Assist Tomogr 5: 136–141. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[28] Intranazális ingerek intermodális integrációja: Funkcionális mágneses rezonancia képalkotási vizsgálat. Boyle JA, Frasnelli J, Gerber J, Heinke M, Hummel T (2007): Neuroscience 149: 223–231. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[29] A szagok és az irritáció kemoreceptor modalitásainak kölcsönhatása. Cain WS, Murphy CL (1980): Nature 284: 255–257. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[30] Szagtalanítás bináris keverékekben. Cashion L, Livermore A, Hummel T (2006): Biol Psychol 73: 288–297. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[31] Neuronspecifikus markerek expressziója a vomeronazális neuroepitheliumban hat főemlősfajban. Dennis JC, Smith TD, Bhatnagar KP, Bonar CJ, Burrows AM, Morrison EE (2004): Anat Record 281: 1190–1200. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[32] Emberi agyműködés. Frackowiak RSJ (2004): Amszterdam: Elsevier Academic. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[33] Hány alany alkot egy tanulmányt? Friston KJ, Holmes AP, Worsley KJ (1999): Neuroimage 10: 1–5. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[34] Csökkent trigeminális érzékenység anozmiában. Gudziol H, Schubert M, Hummel T (2001): ORL J Otorhinolaryngol Relat Spec 63: 72–75. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[35] A szopás feromonális felszabadulása nyulakban nem függ a vomeronazális szervtől. Hudson R, Distel H (1986): Physiol Behav 37: 123–128. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[36] A szaglási funkció felmérése. Hummel T, Welge‐Luessen A (2006): Adv Otorhinolaryngol 63: 84–98. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[37] A szaglási funkció elvesztése a trigeminális ideg érzékenységének csökkenéséhez vezet. Hummel T, Barz S, Lotsch J, Roscher S, Kettenmann B, Kobal G (1996): Chem Senses 21: 75–79. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[38] A szaglási funkció, az életkor és a nem hatása a trigeminálisan közvetített érzésekre: Kemoszenzoros ingerek lateralizációján alapuló tanulmány. Hummel T, Futschik T, Frasnelli J, Huttenbrink KB (2003): Toxicol Lett 140/141: 273–280. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[39] Intranazális trigeminális funkció ép szaglással és anélkül rendelkező alanyokban. Iannilli E, Gerber J, Frasnelli J, Hummel T (2007): Brain Res 1139: 235–244. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[40] A szteroid kemoszignálok pszichológiai állapotra és hangulatra gyakorolt hatásai nőknél és férfiaknál. Jacob S, McClintock MK (2000): Horm Behav 37: 57–78. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[41] A szteroid kemoszignálok kontextusfüggő hatásai az emberi fiziológiára és hangulatra. Jacob S, Hayreh DJ, McClintock MK (2001): Physiol Behav 74: 15–27. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[42] Új kifejezés a biológiailag aktív anyagok egy osztályára. Karlson P, Luscher M (1959): Feromonok: Nature 183: 55–56. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[43] A fő szaglóhám pusztulása csökkenti a szexuális viselkedést és szaglásvizsgálatát. Keller M, Douhard Q, Baum MJ, Bakker J (2006): Chem Sens 31: 315–323. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[44] Genomikus imprinting és a nemi különbségek evolúciója emlősök reprodukciós stratégiáiban. Keverne EB (2007): Adv Genet 59: 217–243. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[45] A kemoszenzoros receptorok második osztálya a szaglóhámban. Liberles SD, Buck LB (2006): Nature 442: 645–650. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[46] A társasági illatanyagok küszöbérték alatti mennyisége befolyásolja a hangulatot, de nem a viselkedést heteroszexuális nőknél, férfi, de nem női kísérletező által tesztelve. Lundstrom JN, Olsson MJ (2005): Biol Psychol 70: 197–204. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[47] Szenzoros feldolgozás a fő és a járulékos szaglórendszerben: Összehasonlítások és kontrasztok. Meredith M (1991): J Steroid Biochem Mol Biol 39: 601–614. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[48] Az emberi vomeronazális rendszer. Áttekintés. Monti-Bloch L, Jennings-White C, Berliner DL (1998): Ann NY Acad Sci 855: 373–389. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[49] Illékony és nem illékony kémiai jelek hatása a férfi szexuális viselkedésre, melyeket a fő és a járulékos szaglórendszer közvetít. O’Connell RJ, Meredith M (1984): Behav Neurosci 98: 1083–1093. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[50] Hormonális és viselkedési válaszok szagokra: A szaglás, a vomeronazális rendszer és a szexuális élmény szerepe. Pfeiffer CA, Johnston RE (1994): Physiol Behav 55: 129–138. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[51] A férfi hónaljkivonatok feromonokat tartalmaznak, amelyek befolyásolják a luteinizáló hormon pulzáló szekrécióját és a hangulatot a kezelt nőknél. Preti G, Wysocki CJ, Barnhart KT, Sondheimer SJ, Leyden JJ (2003): Biol Reprod 68: 2107–2113. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[52] Egy feromonreceptor gén, amely az emberi szaglónyálkahártyában expresszálódik. Rodriguez I, Greer CA, Mok MY, Mombaerts P (2000): Nat Genet 26: 18–19. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[53] Szaglóanyagok agyi aktivációjának képalkotása emberekben. Savic I (2002): Curr Opin Neurobiol 12: 455–461. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[54] A szaglási funkciókat párhuzamos és hierarchikus feldolgozás közvetíti. Savic I, Gulyas B, Larsson M, Roland P (2000): Neuron 26: 735–745. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[55] A szagos nemi hormonszerű vegyületek szaglása nemenként eltérő hipotalamusz aktivációkat okoz emberekben. Savic I, Berglund H, Gulyas B, Roland P (2001): Neuron 31: 661–668. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[56] Az agyi aktiváció szaglóanyag általi differenciált mintázata: Az aceton és a vanillin összehasonlítása. Savic I, Gulyás B, Berglund H (2002): Hum Brain Mapp 17: 17–27. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[57] Agyi válasz feromonokra homoszexuális férfiaknál. Savic I, Berglund H, Lindstrom P (2005): Proc Natl Acad Sci USA 102: 7356–7361. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[58] Mukociliáris funkció orrpolipban szenvedő betegeknél. Slater A, Smallman LA, Logan AC, Drake-Lee AB (1996): Clin Otolaryngol Allied Sci 21: 343–347. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[59] Vak szaglás: Egy nem észlelt, levegőben terjedő vegyi anyag által kiváltott agyi aktiváció. Sobel N, Prabhakaran V, Hartley CA, Desmond JE, Glover GH, Sullivan EV, Gabrieli JD (1999): Brain 122 (2. rész): 209–217. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[60] Az androsztenol priming feromon lokális átvitelének humorális útvonala az orrüregből az agyba és az agyalapi mirigybe. Stefanczyk‐Krzymowska S, Krzymowski T, Grzegorzewski W, Ws B, Skipor J (2000): Exp Physiol 85: 801–809. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[61] Az ovuláció szabályozása emberi feromonok által. Stern K, McClintock MK (1998): Nature 392: 177–179. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[62] A vomeronazális üreg felnőtt emberekben. Trotier D, Eloit C, Wassef M, Talmain G, Bensimon JL, Doving KB, Ferrand J (2000): Chem Senses 25: 369–380. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[63] A vomeronazális hám kemoszenzoros természetéről felnőtt emberekben. Witt M, Georgiewa B, Knecht M, Hummel T (2002): Histochem Cell Biol 117: 493–509. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[64] A nemi hormonok és a nem hatása a görények illékony anális illatmirigyeinek vonzási küszöbértékeire. Woodley SK, Baum MJ (2003): Horm Behav 44: 110–118. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[65] A fő szaglóhagymában található glomerulusok eltérő aktiválása, anális illatmirigyeinek szagai által: Korai lépés a párazonosításban. Woodley SK, Baum MJ (2004): Eur J Neurosci 20: 1025–1032. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[66] ZuFeromon kimutatás emlősök vomeronasalis neuronjaival. fall F, Kelliher KR, Leiders‐Zufall T (2002): Microsc Res Tech 58: 251–260. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[67] A testszagok funkcionális neuronális feldolgozása eltér a hasonló gyakori szagok feldolgozásától – McGill Egyetem, Montreal // Functional neuronal processing of body odors differs from that of similar common odors., Lundström JN, Boyle JA, Zatorre RJ, Jones-Gotman M., Department of Psychology, McGill University, Montreal, QC, H3A1B1, Canada. National Library of Medicine – National Center for Biotechnology Information (NIH)
[68] Emberi feromonok és a szexuális vonzalom - Human pheromones and sexual attraction. Karl Grammer, Bernhard Fink, Nick Neave Ludwig-Boltzmann-Institute for Urban Ethology, c/o Institute of Anthropology, University of Vienna, Austria. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[69] Kozmetikai parfümök vs. emberi feromonok (természetes kémiai illatok) az emberi nők és férfiak szexuális viselkedésének jelzésében és végrehajtásában - Cosmetic perfumes vs. human pheromones (natural chemical scents) of the human female and male in signalling and performing context of their sexual behaviour. PMID: 19750984 M Zaviacic, V Sisovsky, T Zaviacic: Department of Pathology, Faculty ofMedicine, Comenius University, Bratislava, Slovakia. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[70] A Humanae Vitae és a természetjog tudományos alapjai: Az emberi feromonok szerepe az emberi szexuális viselkedési preferenciákban szájon át szedhető fogamzásgátlók alkalmazásával és a szájon át szedhető fogamzásgátlók vetélést okozó hatásai - A Scientific Basis for Humanae Vitae and Natural Law: The Role of Human Pheromones on Human Sexual Behavior Preferences by Oral Contraceptives and the Abortifacient Effects of Oral Contraceptives. Linacre Q. 2018 May;85(2):148-154. doi: 10.1177/0024363918756191. Epub 2018 Apr 12. PMID: 30046193; PMCID: PMC6056803. Angela Lanfranchi: Breast Cancer Prevention Institute, Whitehouse Station, NJ, USA. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[71] Az emberi test illatai: befolyásolják-e a viselkedésünket? - Human body scents: do they influence our behavior? Mildner S, Buchbauer G. Human body scents: do they influence our behavior? Nat Prod Commun. 2013 Nov;8(11):1651-62. PMID: 24427964. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)
[72] Szaglás, feromonok és élet - Olfaction, pheromones and life. Hoskison EE. Olfaction, pheromones and life. J Laryngol Otol. 2013 Dec;127(12):1156-9. doi: 10.1017/S0022215113002545. Epub 2013 Nov 26. PMID: 24280062. Országos Orvostudományi Könyvtár - PubMed – Nemzeti Biotechnológiai Információs Központ (NIH) – National Library of Medicine – National Center for Biotechnology Information (NIH)